Fiksacija ugljenika

Fiksacija ugljenika ili ugljenična asimilacija je proces konverzije neorganskog ugljenika (ugljen-dioksida) do organskih jedinjenja posredstvom živih organizama. Najprominentniji primer je fotosinteza. Hemosinteza je još jedna forma ugljenične fiksacije, do koje može doći u odsustvu sunčevog svetla. Organizmi koji rastu putem fiksacije ugljenika se nazivaju autotrofima. Autotrofi obuhvataju fotoautotrofe, koji sintetišu organska jedinjenja koristeći energiju sunčevog svetla, i litoautotrofe, koji sintetišu organska jedinjenja koristeći energiju neorganske oksidacije. Heterotrofi su organizmi koji rastu koristeći ugljenik koji su fiksirali autotrofi. Heterotrofi koriste organska jedinjenja kao izvor energije i za izgradnju telesnih struktura. „Fiksirani ugljenik”, „redukovani ugljenik”, i „organski ugljenik” su ekvivalentni termini za razna organska jedinjenja.[1]
Neto vs bruto CO2 fiksacija

Procenjuje se da se približno 258 milijardi tona ugljen-dioksida fotosintetički konvertuje svake godine. Najveći deo fiksacije se odvija u morskom okruženju, a posebno u oblastima bogatim nutrijentima. Bruto količina fiksiranog ugljen-dioksida je znatno veća pošto približno 40% se respiratorno konzumira nakon fotosinthesze.[1] Imajući u vidu veličinu ovog procesa, razumljivo je da je ribuloza-bisfosfatna karboksilaza najzastupljeniji protein na Zemlji.
Pregled puteva
Šest autotrofnih puteva ugljenične fiksacije je poznato, prema podacima iz 2011. godine. Kalvinov ciklus fiksira ugljenik u hloroplastima biljki i algi, i u cijanobakterijama. Na taj način se isto tako fiksira ugljenik u beskiseoničnog fotosintezi u jednom tipu proteobakterija zvanih purpurne bakterije, i u nekim nefototrofnim proteobakterijama.[2]
Kiseonična fotosinteza
U fotosintezi, energija iz sunčevog svetla pokreće put fiksacije ugljenika. Kiseoničnu fotosintezu koriste primarni proizvođači— biljke, alge, i modrozelene bakterije. Oni sadrže pigment hlorofil, i koriste Kalvinov ciklus da autotrofno fiksiraju ugljenik. Taj proces se odvija na sledeći način:
- 2H2O → 4e− + 4H+ + O2
- CO2 + 4e− + 4H+ → CH2O + H2O
U provom koraku, voda se disocira u elektrone, protone, i slobodni kiseonik. Time se omogućava upotreba vode, jedne od najzastupljenijih supstanci na Zamlji, kao elektronskog donora. Oslobađanje slobodnog kiseonika je sporedni efekat od enormnih konsekvenci. Prvi korak koristi energiju sunčeve svetlosti za oksidovanje vode do O2, i ultimatno za produkciju ATP molekula
- ADP + Pi ⇌ ATP + H2O
i reduktanta, NADPH
- NADP+ + 2e− + 2H+ ⇌ NADPH + H+
U drugom koraku, zvanom Kalvinov ciklus, odvoja se stvarna fiksacija ugljenika. Ovaj proces konzumira ATP i NADPH. Kalvinov ciklus u biljkama je dominantni vid ugljenične fiksacije na kopnu. Isti pristup je zastupljen u algama i modrozelenim bakterijama, u kojima se odvija najveći deo ugljenične fiksacije u okeanima. Kalvinov ciklus konvertuje ugljen-dioksid u šećer, kao triozni fosfat (TP), koji je gliceraldehid 3-fosfat (GAP) zajedno sa dihidroksiaceton fosfatom (DHAP):
- 3 CO2 + 12 e− + 12 H+ + Pi → TP + 4 H2O
Jedna alternativna perspektiva sumira NADPH (izvor e−) i ATP:
- 3 CO2 + 6 NADPH + 6 H+ + 9 ATP + 5 H2O → TP + 6 NADP+ + 9 ADP + 8 Pi
Formula za neorganski fosfat (Pi) je HOPO32− + 2H+. Formule za triozu i TP su C2H3O2-CH2OH and C2H3O2-CH2OPO32− + 2H+
Reference
- ^ а б Geider, R. J., et al., "Primary productivity of planet earth: biological determinants and physical constraints in terrestrial and aquatic habitats", Global Change Biol. 2001, 7, 849–882. . doi:10.1046/j.1365-2486.2001.00448.x. Недостаје или је празан параметар
|title=(помоћ) - ^ Swan BK, Martinez-Garcia M, Preston CM, Sczyrba A, Woyke T, Lamy D, Reinthaler T, Poulton NJ, Masland ED, Gomez ML, Sieracki ME, DeLong EF, Herndl GJ, Stepanauskas R (2011). „Potential for chemolithoautotrophy among ubiquitous bacteria lineages in the dark ocean”. Science. 333 (6047): 1296—300. Bibcode:2011Sci...333.1296S. PMID 21885783. doi:10.1126/science.1203690.
Literatura
- Berg IA (2011). „Ecological aspects of the distribution of different autotrophic CO2 fixation pathways”. Appl. Environ. Microbiol. 77 (6): 1925—36. PMC 3067309
 . PMID 21216907. doi:10.1128/AEM.02473-10.
. PMID 21216907. doi:10.1128/AEM.02473-10. - Keeling PJ (2004). „Diversity and evolutionary history of plastids and their hosts”. Am. J. Bot. 91 (10): 1481—93. PMID 21652304. doi:10.3732/ajb.91.10.1481.
- PJ, Keeling (2009). „Chromalveolates and the evolution of plastids by secondary endosymbiosis”. J. Eukaryot. Microbiol. 56 (1): 1—8. PMID 19335769. doi:10.1111/j.1550-7408.2008.00371.x.
|access-date=захтева|url=(помоћ) - Keeling PJ (2010). „The endosymbiotic origin, diversification and fate of plastids”. Philos. Trans. R. Soc. Lond. B Biol. Sci. 365 (1541): 729—48. PMC 2817223 . PMID 20124341. doi:10.1098/rstb.2009.0103.
- Timme RE, Bachvaroff TR, Delwiche CF (2012). „Broad phylogenomic sampling and the sister lineage of land plants”. PLoS ONE. 7 (1): e29696. Bibcode:2012PLoSO...7E9696T. PMC 3258253 . PMID 22253761. doi:10.1371/journal.pone.0029696.
- Spiegel FW (2012). „Evolution. Contemplating the first Plantae”. Science. 335 (6070): 809—10. Bibcode:2012Sci...335..809S. PMID 22344435. doi:10.1126/science.1218515.
- Price DC, Chan CX, Yoon HS, Yang EC, Qiu H, Weber AP, Schwacke R, Gross J, Blouin NA, Lane C, Reyes-Prieto A, Durnford DG, Neilson JA, Lang BF, Burger G, Steiner JM, Löffelhardt W, Meuser JE, Posewitz MC, Ball S, Arias MC, Henrissat B, Coutinho PM, Rensing SA, Symeonidi A, Doddapaneni H, Green BR, Rajah VD, Boore J, Bhattacharya D (2012). „Cyanophora paradoxa genome elucidates origin of photosynthesis in algae and plants” (PDF). Science. 335 (6070): 843—7. Bibcode:2012Sci...335..843P. PMID 22344442. doi:10.1126/science.1213561. Архивирано из оригинала (PDF) 14. 05. 2013. г. Приступљено 10. 4. 2012.

- p
- r
- u
| Aerobno disanje | |
|---|---|
| Anaerobno disanje |
|
| Proteinski metabolizam | |||||
|---|---|---|---|---|---|
| Metabolizam ugljenih hidrata (katabolizam ugljenih hidrata i anabolizam) |
| ||||
| Lipidni metabolizam (lipoliza, lipogeneza) |
| ||||
| Aminokiselina | |||||
| Nukleotidni metabolizam | |||||
| Drugi |
Mapa metabolizma | |||||||
|---|---|---|---|---|---|---|---|
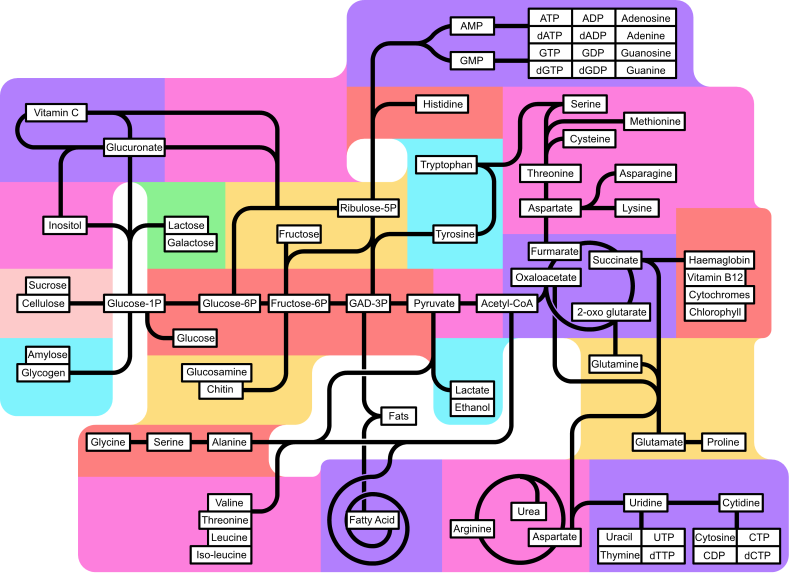 Metabolizam glukoronata Inter-konverzija pentoze Metabolizam inozitola Metabolizam celuloze i saharoze Metabolizam skroba i glikogena Metabolizam drugih šećera Pentozo-fosfatni put Metabolizam amino šećera Sinteza malih aminokiselina Sinteza razgranatih aminokiselina Biosinteza purina Metabolizam histidina Sinteza aromatičnih aminokiselina Piruvat dekarboksilacija Fermentacija Metabolizam masnih kiselina Sinteza aspartatne amino kiselinske grupe Metabolizam porfirina i korinoida Sinteza glutamat amino kiselinskih grupa
| |||||||
| |||||||


Portali:
 Molekularna i ćelijska biologija
Molekularna i ćelijska biologija Hemija
Hemija Biologija
Biologija
| Нормативна контрола |
|
|---|










